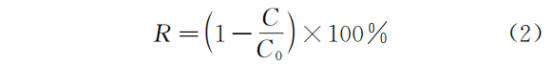揭秘全程氨氧化菌:从其发现、生态分布到生理生化特性
早在1890 年,俄国微生物学家Winogradsky 首次发现并提出了硝化作用,传统生物硝化过程包括亚硝化和硝化两个生化反应。亚硝化过程是在氨氧化菌(ammonia oxidizing bacteria,AOB)的作用下,将氨氮氧化为亚硝态氮[式(1)]。硝化过程是在亚硝酸盐氧化菌(nitrite oxidizing bacteria,NOB)的作用下,将亚硝态氮氧化为硝态氮[式(2)]。因此,硝化过程由AOB和NOB 两类菌群协同作用完成(图1)。然而,2006 年Costa 等根据分子动力学的最佳反应路径长度理论,推测硝化反应可由一种微生物催化完成,并将这类微生物命名为全程氨氧化菌(Comammox)[式(3)]。在2015 年,Daims 等和Kessel 等通过对不同样品的富集和宏基因组测序等技术研究,发现了Comammox的存在。而在2016 年,Pinto 等通过宏基因组测序和荧光原位杂交等技术再次证实了Comammox 的存在。此后,大量学者对Comammox 在自然和人工生态系统中的分布规律以及生理生化特性进行了广泛深入的研究,同时其代谢途径的发现也激发了研究者对其在不同生态系统中的生态学意义的研究。文中详细综述了Comammox 在不同生境中的分布规律及丰度,旨在对其生理生化性能和分布机制有更全面的了解,并揭示Comammox 生态位特征以及对Comammox 的工程实践起到一定的指导意义。


图1 生物硝化过程主要功能微生物和酶
1 Comammox 的发现
Comammox 具备独立将氨氧化为硝酸盐的能力,这一发现颠覆了氨氧化和亚硝酸盐氧化是完全独立两个过程的观点,并重新定义了地球氮循环的关键过程(图2)。

图2 氮循环的主要过程
注:Anammox 为厌氧氨氧化菌; AOA 为氨氧化古菌。
在Costa 等预测后的近10 年内,始终无人证实Comammox 的存在。直到2015 年,Daims 等从石油探井热水管壁上提取生物膜,并经过一系列培养后得到富集培养物,对富集培养物进行宏基因组测序证实了Comammox 的存在,并成功分离出第一株Comammox (Candidatus Nitrospira inopinata, Ca.N.inopinata),这是首次发现Comammox 的存在。同一年,Kessel 等以水产养殖系统厌氧池的生物膜作为接种物进行富集培养,通过分子生物学技术进一步分析发现系统中含有2 种不同的Nitrospira 菌株,并且这两种菌株均含有Comammox amoA 基因,被命名为Candidatus Nitrospira nitrosa(Ca.N.nitrosa)和Candidatus Nitrospira nitrificans(Ca.N.nitrificans)。2016 年,Pinto 等通过16S rRNA 测序和nxrA 基因分析证实了在饮用水系统中存在一种将氨完全氧化为硝酸盐的微生物,并提出NOB 和AOB 与AOA 丰度不平衡的原因可能是Comammox 独立氧化氨氮的能力引起的。2021 年,Sakoula 等在缺氧条件下接种含有两种不同的Comammox 的生物质,获得了一种新的高度富集的Comammox,初步命名为Candidatus Nitrospira kreftii(Ca.N.kreftii)。到目前为止,报道的4 种Comammox 均 属 于clade A (图3)。富 集Comammox 的试验条件如表1 所示。
表1 富集Comammox 的试验条件


图3 Comammox 的分类
2 Comammox 的分布特征
自发现Comammox 后,研究者利用Comammox amoA 基因数据在公开数据库进行比对,发现在土壤、湖泊和沉积物以及饮用水处理厂和污水处理厂等生境中均有Comammox 的分布。并将Comammox 与典型硝化菌在几个生态系统中的相对丰度进行了比较,以阐明其对硝化作用的潜在贡献。
2.1Comammox 在土壤中的分布
Comammox 在土壤中被检测到,但在森林土壤中仅检测到了clade B amoA 基因序列,未检测到clade A,而在水稻土壤和水稻根际中则检测到了clade A和clade B。Hu 等使用Pjevac 设计的引物组进一步检测了5 个森林生物群落采集的300 个样本中Comammox、AOA、AOB 和NOB 的丰度。结果发现,在这些森林土壤中,Comammox 的丰度非常高,且在整个pH 梯度上比AOB 和NOB 更为突出。
2.2Comammox 在湖泊沉积物中的分布
Ca.N.nitrosa 和Ca.N.inopinata 与富营养化湖泊沉积物中的Comammox 密切相关,Xu 等发现了富营养化湖泊沉积物中的clade A,而有关Comammox中具体含量的报道较少。Comammox amoA 基因普遍存在于河岸带和河流沉积物中。虽然河岸带中Comammox 的丰度低于河流沉积物,但总体而言,Comammox 的丰度普遍高于AOB 的丰度。
2.3Comammox 在饮用水处理系统中的分布
饮用水处理厂的出水是典型的寡营养环境,其中Nitrospira 丰度非常高。在快速砂滤池的微生物中,Nitrospira 是优势菌属,其丰度比Nitrobacter 高4 个数量级,比AOB 高出近2 个数量级,因此研究者推测饮用水处理系统可能存在Comammox。研究者在饮用水系统中发现了氨被氧化为硝酸盐的宏基因组证据,证明饮用水处理系统中存在大量的Comammox。研究发现,在所研究的12 个饮用水处理厂的砂滤池中均含有丰富的Comammox,且数量超过了AOB 和AOA,成为主要的氨氧化微生物。这些生境可能有利于微生物群落的附着和Comammox 的生长而导致Comammox 丰度较高。
2.4Comammox 在污水处理系统中的分布
由于污水处理的过程主要依赖于细菌的作用,污水处理系统中细菌群落的功能和结构对提高处理效率非常重要。在污水处理厂中,某些细菌容易富集在氧浓度和氨氮浓度逐渐变化的生物膜上,这种环境变化非常适合Comammox 的生长。因此,Comammox 可能广泛存在于污水处理厂中并在硝化过程中起重要作用。
Chao 等的研究发现,在活性污泥和生物膜中都存在Nitrospira, 并含有amoA 基因, 这为Comammox 的存在提供了宏基因组证据,因此,推测污水处理厂中存在Comammox。研究者 利用qPCR 引物对污水处理厂活性污泥中的微生物进行定量分析,发现硝化菌和反硝化菌丰度稳定,并且存在Comammox。通过新设计的Comammox amoA 引物对污水处理厂的活性污泥样本进行检测,发现Comammox amoA 的丰度是AOB amoA 的184 倍,且Ca.N.nitrosa 占Comammox 的94.3%。Annavajhala等对来自美国、丹麦和新加坡的大型污水处理厂的样本进行了宏基因组分析,结果发现Comammox基因序列始终占总基因组的0.28%~0.64%。在对荷兰和中国的6 个活性污泥系统的细菌群落进行研究时,发现大多数系统都有Comammox 存在,但相对丰度较低。然而,一些研究表明在污水处理厂中Comammox 的丰度较高,Beach 等设计了3 组Comammox amoA 的特异性引物,用于检测活性污泥中的Comammox,结果显示,在相对低DO 条件下,Ca.N.nitrosa 在活性污泥中的丰度较高,并且在污泥停留时间较长的反应器中丰度最高。同样,在富含有机碳的硝化-反硝化系统中,即使有AOB 存在的情况下,Comammox 仍然是优势菌群。研究人员发现,Comammox 在污水处理厂中非常活跃,并具有惊人的高代谢潜力。与 AOM 相比,Comammox 表现出高度的分类和多样性,并且其高丰度可能归因于Nitrospira 的多功能代谢,例如利用尿素、氢和甲酸等获取能量。
从以上的讨论可以看出,研究者们在活性污泥中检测到大量的Comammox amoA 基因,这表明Comammox 在污水处理系统中广泛存在并承担了重要的作用。这些发现清楚地表明,与其他硝化菌相比,Comammox 在污水处理厂中更具竞争优势,意味着Comammox 在工程应用方面具有广阔的前景(表2)。此外,还需要进一步研究Comammox 在污水处理系统中的代谢差异和生态位分异,为污水处理脱氮工艺提供新的思路和方向。
表2 Comammox 与典型硝化菌的比较

3 结论
研究者们对Comammox 富集培养物进行生理生化试验分析以及对比不同环境系统中的基因组数据,初步了解Comammox 的生理生化特性(表2)。
3.1代谢多样性
Comammox 的代谢具有多样性,除了利用无机氮作为底物外,还可以利用简单的无机或有机物进行代谢。基因组分析显示,在已知的Comammox 中,存在编码尿素酶蛋白和相关三磷酸腺苷结合盒转运体(ABC)运输系统的基因,这使得尿素可以替代氨氮进行生物合成。Palomo 等从快速砂滤池中提取的Comammox 基因组中发现了多种尿素转运蛋白,包括高亲和力尿素ABC 转运蛋白、尿素羧化相关转运蛋白和尿素的膜外孔蛋白。这一特性可能使Comammox 能够在尿素浓度较低或波动较大的环境中富集生存,如肥沃的土壤、污水处理厂以及许多水生系统。除了降解尿素,Comammox 还能够代谢其他物质,比如,Ca.N.inopinata 还具有代谢氧化氢和硫化物的能力。另外,编码甲酸脱氢酶的基因在Comammox clade B 被发现,而在clade A 中没有发现,这为甲酸在缺氧过渡带中进行生态位分离提供了机会。在进行基因组分析时发现,氰酸盐水合酶基因仅在 NOB 中被检测到, 在Comammox 中并未检测到,这表明在含氰酸盐的环境中NOB 可能比Comammox 更具有优势。此外从厌氧池中获得的Comammox 编码并转录了醋酸盐发酵的基因,这在NOB 中是意想不到的。令人惊讶的是,该菌株还编码了一种硝酸还原酶,这种酶在任何其他NOB 中尚未发现,这可能使该菌株在缺氧条件下比其他NOB 具有竞争优势。
3.2寡营养环境中更具有竞争力
研究人员发现,Comammox 在氨氮浓度较低的寡营养条件下,相比于富营养环境,更具竞争力。以往的研究认为,在氨氮浓度较低的环境中AOA 更具竞争优势,因为其的氨氮半饱和常数普遍低于AOB。然而,比较AOA、AOB 和Comammox 的动力学常数后发现,在寡营养环境中Comammox 表现出更强的竞争力。 Kits 等对Ca.N.inopinata进行动力学分析发现:(1)Ca.N.inopinata 的氨氮半饱和常数Km(app)= 63±10 nmol/L,低于大多数的AOA 和AOB 的氨氮半饱和常数;(2)Comammox氧化每摩尔氨氮的生长率比 N.gargensis 和N.viennensis 分别高32.3%和29.7%,这可能是因为Ca.N.inopinata 在硝化过程中能够获得更多能量。 表明Comammox 像AOA 一样能够在氨氮浓度较低的环境中生长并占据优势。Sakoula 等富集到的Ca.N.krefti 与Ca.N.inopinata 相似,也具有较高的氨氮亲和力[Km(app)≈0.040±0.01 μmol/L]和较高的亚硝酸盐亲和力[Km(app) = 12.5±4.0 μmol/L],说明了Comammox 可能更适应营养条件有限的环境。
Comammox 的基因组中含有碱性磷酸酶,而AOA 和AOB 不含,Comammox 可以在磷限制条件下高表达,因此Comammox 在磷含量较低的环境中具有竞争潜力。与典型NOB 不同,Comammox 基因组含有铜稳态基因(cop CD 和cop AB),并与β-AOB 同源物的序列高度相似。这些蛋白质具有较高的Cu2+耐受性,这可能使Comammox 能够在Cu2+较低的环境中存活。这两方面都非常重要,因为缺乏磷和铜都会影响工程环境中的硝化过程。综上所述,全程硝化菌可能更适应营养条件有限的环境。
3.3较高的DO 亲和力
目前的理论和研究表明Comammox 适应于低DO 环境。与Nitrospira 中其他细菌一样,Comammox具有编码rTCA 的基因。在电子传递方面也揭示了Comammox 适应于微氧环境,它含有的细胞色素bd 是一种在电子传递链中发挥作用的氧化还原酶,对O2 有较高的亲和力并可以在氧浓度较低的环境下表达,因此该酶可能使Comammox 在低DO 环境中具有优势。此外,Comammox 还含有fnr 型调控蛋白,在其他微生物中fnr 在厌氧或微氧条件下作为氧气传感器参与细胞代谢的基因调节和信号传递。虽然Comammox 体内此类基因和蛋白质的具体功能尚未确定,但Camejo 等预测该因子可以调控Comammox 中类似的基因。在试验研究中,Kessel 等在低氧环境中富集厌氧氨氧化细菌时,发现了厌氧氨氧化菌和Comammox 的共存物,这表明了Comammox 可以在低DO 环境中生存。Camejo等在微好氧条件下运行序批式活性污泥反应器(SBR)并成功富集到了Comammox,且在其基因组中发现了与微氧代谢相关的基因。
因此,综合以上分析可以得出结论,Comammox在低DO 的条件下更容易生长和富集。相较于AOA和AOB,Comammox 对氧气的亲和力更高,这意味着在实际的工程应用中,以Comammox 为主的硝化工艺可以在低DO 条件下替代传统的高DO 硝化工艺,从而节省能源。
4 展望
Comammox 的发现和广泛环境分布,对于深入了解完全硝化过程调控机制具有重要意义。 自2015 年发现以来,已证实Comammox 在自然和人工生态系统中广泛存在,并在氮素转化中扮演重要角色。 相比于传统硝化菌,Comammox 展示了更高的氨氮亲和力和相对较低的氨氮氧化率,使其能够在高度寡营养的环境中生存。 然而,关于Comammox的富集培养、理化性质、对全球氮循环的贡献等问题尚未得出定论。 需要进一步了解Comammox 的生态位专门化、种群动态、代谢多功能性和系统发育结构以及与其他微生物的相互作用,才能得出关于Comammox 在生态学意义上的一般性结论。 研究人员可以从以下方面进行探索。
(1)目前为止,研究人员只分离出一种纯培养物Ca.N.inopinata,尚未从陆地生境中获得纯种Comammox 培养物。 因此,研究人员需进一步采用富集培养技术,对不同生境下的Comammox 进行富集培养,以分离出高纯度的Comammox 株。 通过对这些菌株进行基因表达、增殖代谢和环境因素等之间的关系分析,可以在分子细胞水平上探究Comammox 物种进化演替的规律,了解其能量摄取来源,以及与其他功能微生物之间的协同竞争机制。
(2)通过研究硝化菌的物种丰度和群落结构特征,以及分析 Comammox、AOB 和AOA 的氨氧化速率,研究Comammox 和NOB 的亚硝酸盐氧化速率,可以进一步提高对氮循环微生物家族的认识,深入研究氮元素在生物地球循环中的作用。
(3)完全氨氧化过程被认为是一种“绿色过程”,已证实Comammox 含有产生N2O 的相关酶和基因,并可能通过NH2OH 转化产生N2O,但由于基因组中缺乏一氧化氮还原酶,Comammox 在硝化过程中释放的N2O 远低于AOB,这意味着Comammox不是N2O 的主要来源。 因此,Comammox 对全球气候变化的贡献仍需进一步探讨。
(4)探究抑制Comammox 进行硝化反应的因素也十分重要。 一项研究表明,氯酸盐可能是Comammox 进行硝化的潜在抑制剂,因为它可以被硝酸还原酶降解。 不过,氯酸盐对硝酸还原酶是否有抑制作用还需要进一步证实。
(5)Comammox 可以与厌氧氨氮氧化菌共生并为其提供NO2-,这种组合能够降低曝气要求,从而在处理系统中为硝化作用带来操作上的优势,为研发能量和碳源要求更少的脱氮工艺开辟了希望。然而,目前Comammox 工艺的设计参数仍然未知。因此,要全面推行Comammox 工艺还需要进行更广泛的研究。
(6)应广泛开展以反应器为中心的研究,重点确定边界条件,这有助于实现Comammox 的早期大规模应用。 在自然和工程系统中,Comammox 与其他细菌群的相互作用仍然有限。 因此,在未来探索Comammox 与其他细菌之间的相互作用,也有助于评价Comammox 的反应器适应性。
通过深入研究Comammox 的生态学和生理学特征,将更好地认识氮循环和生态系统功能。 同时,还需要进一步探索Comammox 的生态适应机制、群落结构以及其在全球氮循环中的作用,以便更好地管理和利用氮资源,保护生态环境。
作者简介
李晓丽(1999— ),女,硕士,研究方向为全程氨氧化菌的富集培养及其动力学特性,E-mail: lxl19991209@163.com。
通信作者:孙洪伟,男,教授,E-mail:sunhw@ytu.edu.cn。