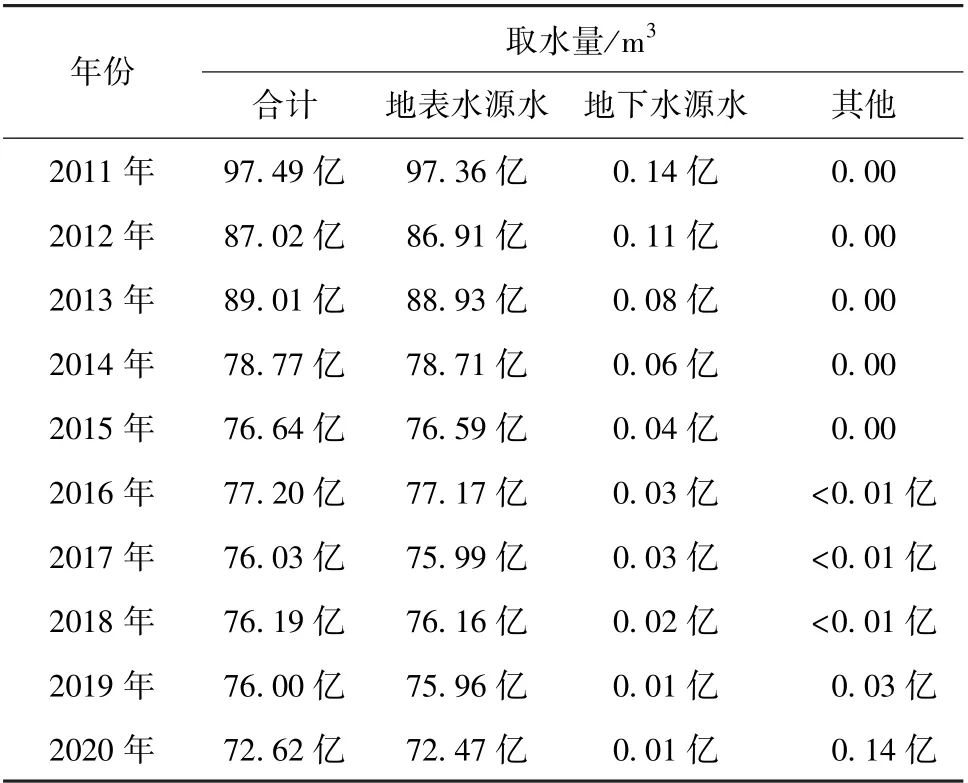
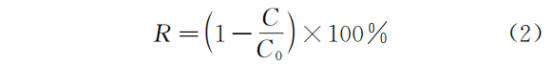学术回溯 | 模拟评价由捕食和病毒引起的污泥减量
编者按:往期内容介绍了对细菌内源衰减特征相关探究,主要包含细胞死亡、活性降低等。本期就内源衰减另一主要过程——高等微生物捕食与病毒感染进行剖析。文章通过扩展活性污泥2号模型(ASM2d),模拟聚磷菌(PAOs)富集SBR系统中微生物反应,并预测活性污泥系统中高等微生物捕食和病毒感染对污泥减量的影响作用。结果表明,在PAOs富集SBR系统中,捕食是污泥减量的主要原因,其贡献程度大约是病毒感染的两倍。对所选关键参数敏感性分析表明,捕食率和病毒感染率对出水磷浓度恶化情况均具有阈值,分别为0.014 h-1、0.009 h-1。在此条件下,捕食和病毒感染过程对污泥减量最大贡献程度分别为21%和9%。本文发表于《Water Research》(2011年11月)
整理 | 依伙古日
责编 | 郝晓地
文章亮点
01、使用扩展ASM2d模型同时评估了高等微生物捕食和病毒感染对污泥减量的贡献作用;
02、捕食率和病毒感染率对出水磷浓度恶化情况均具有阈值,分别为0.014 h-1、0.009 h-1;
03、病毒感染能够促进PAOs富集系统中常规异养细菌(OHOs)隐性生长,这使其对污泥减量的贡献作用不及高等微生物捕食作用。
01引言
微生物内源过程可分为细胞水平和群落水平两个层面。前者主要涉及内源呼吸,而后者包括高等生物捕食、病毒感染等。其中,内源呼吸已得到广泛研究和应用,如延时曝气工艺(氧化沟)、好氧污泥消化等。然而,对细胞衰减有重要作用的捕食和病毒感染过程却少有定量。且针对其对细胞衰减、污水处理效能(特别是生物脱氮除磷)的作用机制仍缺乏深入研究。然而,活性污泥系统中的相互作用十分复杂。对此,可通过数学模型对其进行剖析和评价。此外,由于对各衰减过程的计算并无区分,衰减系数模糊,导致现有模型无法明确将捕食和病毒感染作用与其他衰减过程进行区分。为了更好地评估捕食和病毒感染对污泥减量的影响,本文向活性污泥2号模型(ASM2d)中引入3个单独的细胞衰减过程,并用于模拟聚磷菌(PAOs)富集SBR系统中的微生物反应,以校验模型。最后,通过拓展模型预测活性污泥系统中高等微生物捕食和病毒感染对污泥减量的影响作用。
02主要材料与方法
2.1 PAOs富集SBR系统
PAOs富集SBR系统工况同《活性污泥中聚磷菌/聚糖原菌内源衰减特征探究》中的设定。系统稳定后以SRT=15 d,进水COD=440 mg/L(30 d以乙酸盐为碳源,15 d以丙酸盐为碳源交替运行)用以模型验证。
2.2 扩展模型的构建
采用ASM2d作为模型框架,并向其引入PAOs的糖原代谢过程。此外,为了同时考虑捕食和病毒感染过程,将模型中的细胞衰减过程分为高等微生物捕食、病毒感染和其他衰减作用3个过程(如有毒物质、细胞自然死亡等),并引入相应的化学计量矩阵和动力学速率表达式(详情请参阅原文)。其中,由于PAOs富集SBR系统中投加了硝化抑制剂,且进水中的NH4+-N仅用作细菌合成代谢(系统中NO2--N/NO3--N<0.1 mg N/L),因此从模型中删除了相关过程(S_NH4)。
由于SBR系统在运行过程中存在体积变化,因此,在AQUASIM中利用2个不同体积的完全混合池交替运行以实现对此过程的模拟(详情请参阅原文)。并采用编程和公式(1)、(2)对曝气过程进行模拟。

式中:t_period为AQUASIM中的公式变量,可以表示为t_period=t mod 6;2.1667和4.8333为曝气开始和结束的时间;S0(t)为时间t时液相中DO浓度;82为氧转移系数;7.95为22℃时O2在液相中的最大溶解度。AQUASIM中SBR配置的示意图如图1所示。

图1 拓展模型校准结果(图片来自原文)
2.3 扩展模型的校验
通过第一组实验结果校准扩展模型。其中,为了简化校准程序,尽可能少地改变参数,仅对6个动力学参数(b_H,b_PAO,b_PHA,b_Gly,b_PP,和q_PP)进行校准,以匹配混合液体挥发性悬浮固体浓度(MLVSS)、PAOs浓度(XPAO)、死菌浓度(X_I)、挥发性脂肪酸浓度(VFA: S_A)、磷浓度(S_PO4)和PAOs中的糖原浓度,如图1所示。此外,在第一组实验后的第90天,利用第二组实验结果验证校准后的拓展模型。结果显示模型能够较好地描述SBR系统性能(如图2所示)。

图2 拓展模型验证结果(图片来自原文)
03结果和讨论
3.1 模型校验
SBR系统的稳定根据系统中MLSS和MLVSS,以及每个工作周期中VFA、磷酸盐和糖原浓度的稳定动态变化判断确定。并且通过寡核苷酸探针测定沉降污泥中的PAOs占比高达细菌总数的91%。
在扩展模型中,捕食者的衰减系数(0.15 h-1)和捕食率(0.01 h-1)直接引自参考文献(已由基于实验室规模的SBR系统模型模拟确定)。此外,根据参考文献,病毒感染系数取为0.006 h-1。在校准和验证之后,可以评估病毒感染系数对出水水质的敏感性(图4B)。经过第一次实验结果与模拟结果校准之后,模拟结果接近实验结果(图1),校准成功。
利用校准的模型和实验的另一操作条件(SRT=15 d和COD=220 mg/L),进行了模型验证,如图2所示。与图1相比,在图2中可以观察到更高的MLVSS和糖原含量。可以说明经校准和验证后的模型可以预测富集PAOs的SBR系统的性能,从而也可以评估捕食和病毒感染对污泥减量化造成的影响。
3.2 捕食率和病毒感染对污泥减量的敏感性分析
首先利用经过校验的拓展模型评价SBR系统中捕食对污泥减量的贡献,模拟结果如表1所示。结果显示,捕食会造成污泥减量18%(剩余污泥量从1140 mg COD/d减少到937 mg COD/d)。并且随着捕食量的增加,活菌数量也会降低(12%)。
表1 存在/不存在捕食的模拟结果(来自原文)

不考虑该负面影响的前提下,进一步模拟以观察捕食率(Ep)对污泥产量和污水中磷浓度的敏感性。结果显示,随着捕食率的增大,剩余污泥量逐渐降低,但在捕食率为0.014 h-1时出水磷浓度显著增加(图3A)。因此,若要保持出水磷浓度,则应控制捕食率小于0.014 h-1(此时污泥减量达21%)。此外,从捕食率对MLVSS等指标的敏感性分析(图3B)进一步说明,出水磷浓度的恶化与捕食率的增大显著相关。这是由于生物除磷系统除了受到SRT的限制以外,还会受到最大捕食率的限制。若超过这个极限,PAOs将从系统中被完全抹除,致使系统除磷效能降低。

图3 捕食率对污泥产量和出水磷浓度(A)、MLVSS、XPAO、X_H(普通异养生物)、X_I(B)的敏感性(图片来自原文)
同理,该模型模拟了分别在涉及和不涉及病毒感染的两种情况下对污泥产量的影响(其他参数保持不变),模拟结果如表2所示,病毒感染使得剩余污泥量减少8%。实际上,这是首次证明病毒感染对污泥最小化的贡献。
表2 涉及/不涉及病毒感染的模拟结果(来自原文)

有趣的是,OHOs(普通异养微生物)的数量随病毒活动的增加而增加。这种现象可以解释为,在PAOs富集系统中,有机底物(COD/VFAs)几乎都被PAOs吸收,并在厌氧条件下形成胞内PHA。因此,OHOs只能通过PAOs和/或PAOs排出的代谢产物(即隐生长)生长。当病毒感染发生时,PAOs死亡并溶出更多的有机产物。这为OHOs提供了更多的生长底物。当然,病毒感染也会导致OHOs的死亡,但显然其生长率高于死亡率。简而言之,高等微生物利用细菌细胞作为底物来产生维持生长,而病毒则利用细菌细胞作为繁殖的宿主。宿主死亡并释放新病毒后,死亡的细胞成为OHOs的生长底物。因此,OHOs数量往往会随着病毒感染增多而增加。
病毒感染率对剩余污泥和出水磷浓度的敏感性分析如图4a所示,随着病毒感染率(Ev)的增加,出水磷浓度增加。但与捕食作用一样,在Ev≥0.009 h-1时出水磷浓度也突然恶化。在这种状态下,剩余污泥量约为920 mg COD/d,相当于污泥产量减少了9%。图4b也进一步说明,由于会导致OHOs的生长,病毒感染对污泥减量的贡献不如高等微生物的捕食作用。

图4 病毒感染率对污泥产量和出水磷浓度(A)、MLVSS、XPAO、X_H(普通异养生物)、X_I(B)的敏感性(图片来自原文)
结论
01通过扩展ASM2d模型,经过校准参数以及模型验证后证实其可以同时评估评估捕食和病毒感染对污泥减量造成的影响。
02敏感性分析表明,捕食率以及病毒感染率都存在阈值,导致捕食作用和病毒感染对污泥减量的影响被限制在21%和9%。
03模拟表明,捕食比病毒感染更有效地控制污泥产量,病毒感染后死亡的细胞会被用于常规异养细菌(OHOs)生长。